— 生態(tài) · 農(nóng)業(yè) · 健康 —

時間:2018-11-14
作者:易科泰
點擊量:
簡介:
TL6000植物熱釋光測量系統(tǒng)
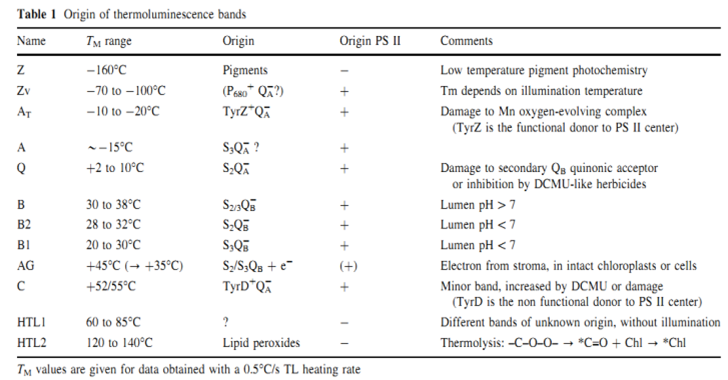
熱釋光(Thermoluminescence,,縮寫TL)是晶體受到輻射照射后,產(chǎn)生了自由電子,。這些電子被晶格缺陷俘獲而積攢起來,,在加熱過程中以光形式釋放出來,。葉綠素熱釋光則是由于活化能壘在生理溫度下限制了諸如電子再結(jié)合等暗反應,因此光化學反應中分離的電子對穩(wěn)定存在于電子載體中。在熱刺激下,, S2QA-,,S2QB- 和 S3QB-電子對的再結(jié)合,使PSII中激發(fā)的單線態(tài)葉綠素分子發(fā)出熱釋光,。然后,,逐漸升高溫度會增加再結(jié)合的比率,從而激發(fā)不同類型的電子對形成TL譜帶,。葉綠素熱釋光能夠揭示光合放氧復合物(OEC)穩(wěn)定性及PSII總體完整性,、QB受體損傷及葉綠體內(nèi)腔 pH值變化等光系統(tǒng) II的深層運轉(zhuǎn)機理。
TL植物熱釋光測量系統(tǒng)是目前唯一商用化的葉綠素熱釋光測量儀器,,針對研究PSII能量水平結(jié)構(gòu)進行了專門設(shè)計,。PSII反應中心光誘導電荷分離導致儲存了吸收光能的激發(fā)電子對的累積。加熱誘導這些激發(fā)電子對的重組,,從而引發(fā)光釋放,,并在一定溫度范圍內(nèi)形成特異性熱釋光曲線。根據(jù)不同釋光曲線的形狀,、峰位和峰值,,可以研究分析關(guān)于特定激發(fā)電子對的能量穩(wěn)定性及PSII反應中心功能等。TL6000為這一系統(tǒng)的最新型號,,最大測量范圍為-100°C到+200°C,,使用范圍更寬,可以對低溫,、高溫段熱釋光進行研究,。

應用領(lǐng)域
1、無損傷測量PSII電子傳遞
2,、PSII對生物/非生物脅迫與結(jié)構(gòu)修飾的適應和應答
3,、脅迫條件下葉綠體內(nèi)能量不平衡的敏感性檢測
4、類囊體膜PSII氧化還原反應
5,、基于峰值溫度轉(zhuǎn)換,,對供體側(cè)與受體側(cè)的氧化還原電位變化進行解釋
6、通過測量熱釋光震蕩模式,,指示S狀態(tài)轉(zhuǎn)換和放氧復合體狀態(tài)
典型樣品
工作原理

熱釋光(Thermoluminescence,,縮寫TL)是晶體受到輻射照射后,會產(chǎn)生自由電子,,這些電子被晶格缺陷俘獲而積攢起來,,在加熱過程中以光形式釋放出來。其基本的實驗過程是將葉片快速冷凍到某一溫度,,之后給葉片一個足夠強,,但時間盡量短(一般<5μs)的單反轉(zhuǎn)光(single turn-over ?ash),用于誘導每個PSII反應中心發(fā)生僅一次的電荷分離,;然后逐漸升溫,,同時測量葉片放出的熱釋光,繪制TL譜帶,。
TL植物光合熱釋光測量系統(tǒng)使用能量足夠強的LED光源,,所釋放5-10μs的方波脈沖能夠飽和所有的PSII反應中心,其溫度控制單元可以在降溫后,,再使樣品的溫度以0.1℃/sec到1.5℃/sec的速率線性增加,。不同的閃光序列及樣品處理能夠使樣品處于不同的能量狀態(tài),不同的溫度下釋放的光能源自光合機構(gòu)的不同結(jié)構(gòu),。分析釋光曲線的形狀,、峰位和峰值,可以研究分析關(guān)于特定激發(fā)電子對的能量穩(wěn)定性及PSII反應中心功能等,。
熱釋光(TL)譜帶的來源及意義
不同型號的控溫方式與范圍

系統(tǒng)組成
TL系列植物光合熱釋光測量系統(tǒng)由3部分組成:主控制分析單元,、外部制冷單元、測量室,。
主控制分析單元:根據(jù)用戶定義方案或軟件內(nèi)置的實驗程序來執(zhí)行實驗過程并采集數(shù)據(jù),。彩色顯示屏可實時顯示測量曲線。
外部制冷單元:
(1)水冷單元(TL 6000/ST標準版配備):包含一個電子控制的抽水泵和內(nèi)部儲水的制冷器,,用于測量室降溫,。
(2)液氮制冷單元(TL 6000/ET溫度擴展版配備):將液氮罐通過管路連接到測量室,通過電子控制的低溫輸出閥可以將測量室溫度控制到最低-100℃,。同時也用于測量結(jié)束后給測量室降溫,。
測量室:包括四個關(guān)鍵組成部分:光源、光電倍增管傳感器,、溫度控制器,、樣品盤。

技術(shù)參數(shù)
?TL 6000/ST標準版:-25℃到+70℃
?TL 6000/ET溫度擴展版:-100℃到+200℃
操作軟件與實驗結(jié)果

典型應用

中科院植物所盧從明研究員是國內(nèi)最早將TL熱釋光技術(shù)用于植物光合研究的科學家之一,,其領(lǐng)導的團隊也一直位于這一研究的國際最前沿,。上圖即為2016年發(fā)表的文獻,通過測量施加DCMU和不同光照條件的葉綠素熱釋光曲線,,評估谷胱甘肽還原酶2對擬南芥PSII維持功能的作用(Ding,,2016)。
產(chǎn)地:歐洲
參考文獻:
1. OHP1, OHP2, and HCF244 form a transient functional complex with the photosystem II reaction center, Y Li, et al, 2019. Plant Physiology 179, 195–208
2. Antimycin A inhibits cytochrome b559-mediated cyclic electron flow within photosystem II, D Takagi, et al, 2019. Photosynthesis Research 139(1–3), 487–498
3. 'Birth defects' of photosystem II make it highly susceptible to photodamage during chloroplast biogenesis, D Shevela, et al, 2019. Physiologia Plantarum 166, 165–180
4. High light acclimation of Chromera velia points to photoprotective NPQ, E Belgio, et al, 2018. Photosynthesis Research 135(1–3), 263–274
5. Comparison of photosynthetic performances of marine picocyanobacteria with different configurations of the oxygen-evolving complex, F Partensky, et al, 2018. Photosynthesis Research 138(1), 57–71
6. Diel regulation of photosynthetic activity in the oceanic unicellular diazotrophic cyanobacterium Crocosphaera watsonii WH8501, T Masuda, et al, 2018. Environmental Microbiology 20(2), 546–560
7. Glutathione reductase 2 maintains the function of photosystem II in Arabidopsis under excess light, S Ding, et al, 2016. Biochimica et BiophysicaActa (BBA) – Bioenergetics1857(6), 665–677
8. Titanium dioxide nanoparticles (100–1000 mg/l) can affect vitamin E response in Arabidopsis thaliana, R Szymańska, et al, 2016. Environmental Pollution 213, 957–965
9. The N-terminal sequence of the extrinsic PsbP protein modulates the redox potential of Cyt b559 in photosystem II, T Nishimura, et al, 2016.Sci Rep. 6, 21490.
10. The PsbY protein of Arabidopsis Photosystem II is important for the redox control of Cytochrome b 559, L von Sydow, et al, 2016. Biochimica et BiophysicaActa (BBA) – Bioenergetics1857 (9),1524–1533

西安研發(fā)中心
微信公眾號

業(yè)務(wù)咨詢
微信號

易科泰公司
微信公眾號

易科泰公司
視頻號
聯(lián)系我們:
地址: 北京市海淀區(qū)高里掌路3號院6號樓1單元101B
電話: 010-82611269/1572
手機: 13671083121
傳真: 010-62465844
Email: [email protected]
友情鏈接:

